2 papers mentioning rco-MIR167a
Open access articles that are associated with the species Ricinus communis
and mention the gene name MIR167a.
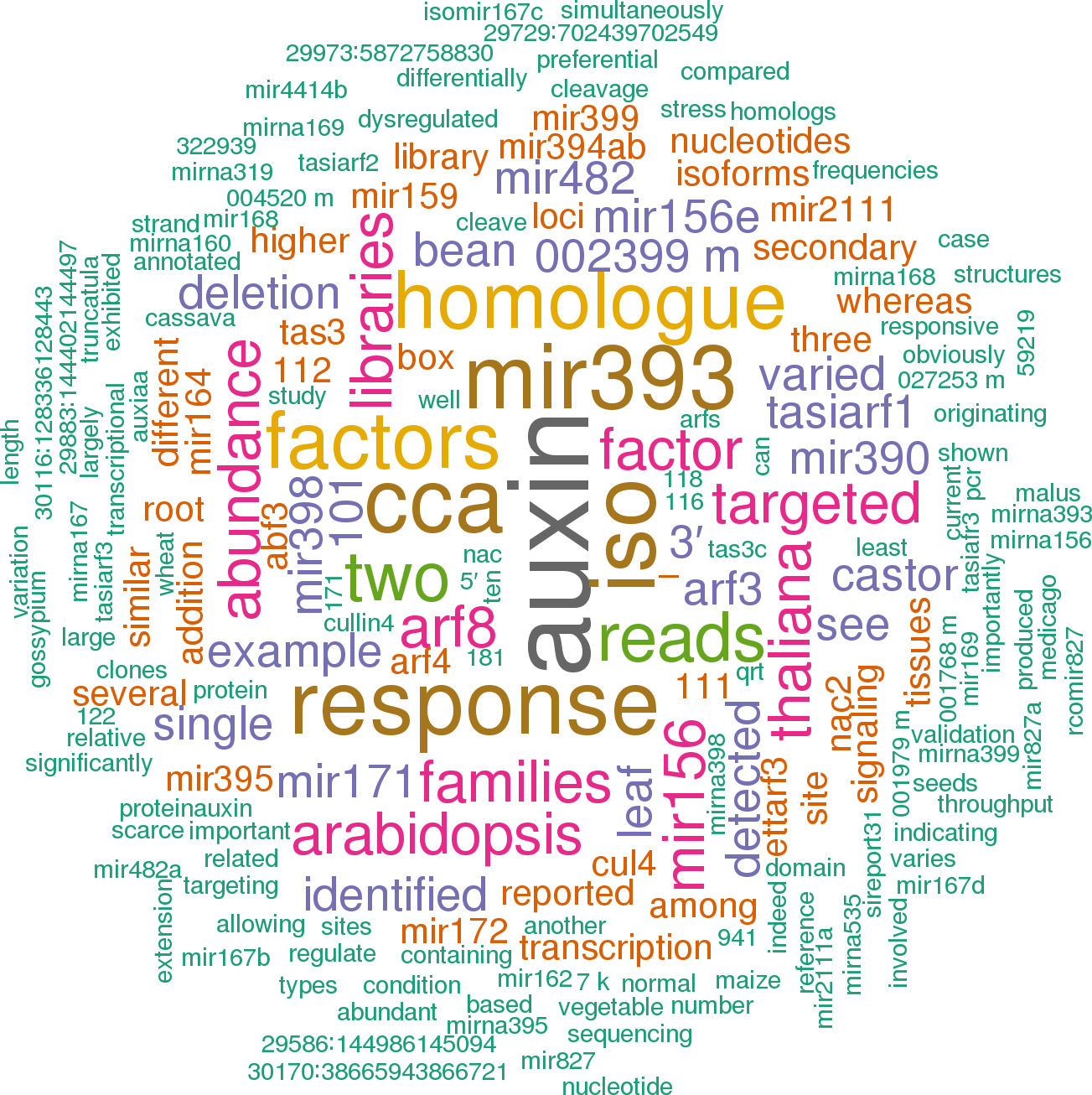
Click the buttons to view sentences that include the gene name, or the word cloud on the right for a summary.
 |
|