Accession
MI0000477
Symbol
HGNC:
MIR146A
Description
Homo sapiens
hsa-mir-146a precursor miRNA
Summary
Caution, this is an AI generated summary based on literature. This may have errors. ?
WARNING: This summary was generated by AI. MIR146A, a microRNA, plays a crucial role in various biological processes, including chondrogenesis, where its absence in knockout MSCs confirmed its function [PMC10110697]. It also exhibits a protective role in diabetic nephropathy and influences lung structure and function through its downstream targets [PMC5354466; PMC10129905].]. MIR146A directly targets STAT1, impacting its expression and activation [PMC9125934], and its expression levels are notably altered in various diseases such as lupus nephritis and esophageal squamous-cell cancer [PMC9013931; PMC6301302].. Variants within the MIR146A gene promoter may affect its expression [PMC9078850], suggesting potential therapeutic applications in MSC-based osteoarthritis therapy [PMC10110697]. Moreover, MIR146A is implicated in the immune response as it is differentially expressed between naïve and memory T cells and is induced by TCR stimulation [PMC4641946]. It also acts as a negative regulator of NFκB by suppressing TRAF6, highlighting its anti-inflammatory potential which could be harnessed using engineered exosomes for therapeutic delivery [PMC6651735; PMC9922687]..
Literature search
749 open access papers mention hsa-mir-146a
(6781 sentences)
(6781 sentences)
Sequence
319175
reads,
1010
reads per million, 137 experiments
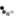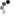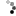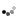
ccgauguguauccucagcuuUGAGAACUGAAUUCCAUGGGUUgugucagugucagaCCUCUGAAAUUCAGUUCUUCAGcugggauaucucugucaucgu
.(((((.((((((.(((((..((((((((((((.((.(((((.((.......))))))).)).)))))))))))).)))))))))))......))))).
.(((((.((((((.(((((..((((((((((((.((.(((((.((.......))))))).)).)))))))))))).)))))))))))......))))).
Structure
c -----u u uU C U g uc cgaug guaucc cagcu GAGAACUGAAUU CA GGGUU ug a ||||| |||||| ||||| |||||||||||| || ||||| || g gcuac uauagg gucGA UUCUUGACUUAA GU UCCag ac u u ugucuc - -C A C - ug
Annotation confidence
High
Do you think this miRNA is real?
Comments
This miRNA sequence is predicted based on homology to a verified miRNA from mouse [1]. Its expression was later verified in human [2,3].
Genome context
chr5: 160485352-160485450 [+]
Disease association
hsa-mir-146a is associated with one or more human diseases in the Human microRNA Disease Database
| Disease | Description | Category | PubMed ID |
|---|
Database links

Mature hsa-miR-146a-5p
| Accession | MIMAT0000449 |
| Description | Homo sapiens hsa-miR-146a-5p mature miRNA |
| Sequence | 21 - UGAGAACUGAAUUCCAUGGGUU - 42 |
| Evidence |
experimental
cloned [2-3] |
| Database links |
|
| Predicted targets |
|
Mature hsa-miR-146a-3p
| Accession | MIMAT0004608 |
| Description | Homo sapiens hsa-miR-146a-3p mature miRNA |
| Sequence | 57 - CCUCUGAAAUUCAGUUCUUCAG - 78 |
| Evidence |
experimental
cloned [3] |
| Database links |
|
| Predicted targets |
|
References
|